×

سلام
ورود و عضویت
همیوغی باکتریایی (Bacterial conjugation) به فرایند انتقال ماده ژنتیک میان سلولهای باکتریایی، از طریق تماس مستقیم سلول به سلول و یا ایجاد ارتباط پل مانند میان دو سلول گفته میشود(۱). همیوغی هم مانند انتقال ژنتیکی (transduction) و ترانسفورماسیون (transformation) روشی از انتقال افقی ژن (horizontal gene transfer) در فرایند انتقال ژن است با این تفاوت که انتقال ژن در این دو نوع(transduction و transformation) از طریق تماس مستقیم سلول به سلول صورت نمی گیرد(۲).
همیوغی باکتریایی اغلب معادل تولید مثل جنسی(sexual reproduction) و یا جفتگیری(mating) در باکتری در نظر گرفته میشود چرا که شامل تبادل مواد ژنتیکی است. البته باید در نظر داشت که این انتقال ماده ژنتیکی، از نوع تولید مثل جنسی نمیباشد چون هیچگونه تبادل گامتی رخ نمیدهد. در فرایند همیوغی، سلول دهنده، عنصر ژنتیکی همیوغی شونده یا متحرک که اغلب یک پلاسمید(plasmid) و یا یک ترانسپوزون(transposon) است را فراهم میکند(۳). اغلب پلاسمیدهای همیوغی شونده دارای سیستمهایی هستند که از طریق آن اطمینان حاصل میکنند که سلول گیرنده دارای عنصر مشابه نباشد. ماده ژنتیکی منتقل شده اغلب برای گیرنده مفید است. این مزایا شامل مقاومت به آنتی بیوتیک(antibiotic resistanse)، تحمل زنوبیوتیک(xenobiotic) یا توانایی استفاده از متابولیتهای جدید است(۴). چنین پلاسمیدهای سودمندی به عنوان همزیستهای باکتریایی (endosymbionts) در نظر گرفته میشوند. عناصر دیگر، میتوانند به عنوان انگل باکتریایی در نظر گرفته شوند که همیوغی در واقع مکانیسم تکامل یافتهای برای گسترش آنها است.
تاریخچه:
پدیده همیوغی در سال ۱۹۴۶ توسط جاشوا لیدربرگ و ادوارد تیتوم کشف شد(۵).


| جاشوا لیدربرگ (سمت راست) و ادورارد لاوری تیتوم (سمت چپ) که پدیده هم یوغی را کشف کردند. این دو بعدها به خاطر ارائه نظریه یک ژن یک انزیم به طور مشترک برنده جایزه نوبل پزشکی در سال 1958 شدند. |
مکانیسم:
مکانیسم عمل همیوغی در تصویر آورده شده است. پلاسمید F(F-plasmid) یک اپیزوم(episome) با طول حدود 100kb میباشد. اپیزوم در واقع پلاسمیدی است که میتواند با استفاده از نوترکیبی همولوگ( homologous recombinant)، خود را درون کروموزوم باکتری وارد کند. پلاسمید F یک مکان آغاز همانندسازی (origin of replication (oriV)) و یک مکان انتقال (oriT) دارد(۳). فقط یک کپی از پلاسمید F میتواند به صورت آزاد و یا به صورت ادغام شده درون کروموزوم باکتری گیرنده وجود داشته باشد. باکتریهای حامل این کپی، F+ (F positive) نامیده میشوند و سلولهای فاقد این پلاسمید، F- (Fminus) نامیده شده و به عنوان سلول گیرنده عمل میکنند.
در میان اطلاعات ژنتیکی، پلاسمید F حامل دو جایگاه کروموزومی tra و trb است که روی هم رفته 33kb طول دارند و شامل ۴۰ ژن میباشند. جایگاه کروموزومی tra شامل ژن pilin و ژنهای تنظیمکننده است که با یکدیگر پیلی(pili) را در سطح سلول شکل میدهند. همچنین این جایگاه شامل ژنهایی است که کدکننده پروتئینهای متصل شونده به سطح باکتریهای F- هستند و آغازکننده هم یوغی میباشند. بر روی مکانیسم دقیق همیوغی شک و تردید وجود دارد و به نظر میرسد که پیلی ساختارهای اصلی انتقال دهنده DNA در حین همیوغی نیستند و به نظر میرسد چندین پروتئین کد شده توسط جایگاههای ذکر شده کانالهایی را بین دو باکتری باز کرده و آنزیم traD که در قاعده پیلی قرار دارد، ادغام دو غشاء را آغاز میکند.
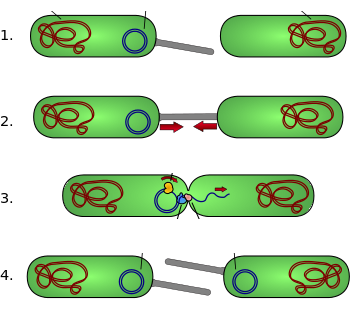
دیاگرام همیوغی: ۱) سلول دهنده، پیلی یا پیلوس تولید میکند. ۲) پیلوس به سلول گیرنده متصل شده و دو سلول را کنار هم میآورد. ۳) پلاسمید دو رشتهای جدا شده و یکDNA تک رشته به سلول دیگر منتقل میشود. ۴) هر دو سلول یک رشته مکمل سنتز کرده تا یک پلاسمید حلقوی دو رشتهای داشته باشند. علاوه بر این هر دو سلول برای خود پیلی تولید کرده تا به عنوان دهنده و یا عامل F (F factor) شناخته می شود.
کاربردهای مهندسی ژنتیک:
همیوغی ابزار مناسبی جهت انتقال مواد ژنتیکی برای اهداف مختلف است. در آزمایشگاه، انتقال موفق از باکتریها به مخمر(۶)، گیاهان، سلولهای پستانداران(۷، ۸) و میتوکندریهای(۹) جدا شده پستانداران گزارش شده است. همیوغی دارای مزایای بیشتری نسبت به انواع دیگر انتقال مواد ژنتیکی میباشد از جمله اینکه باعث ایجاد حداقل اختلال در پوشش غشایی سلول هدف شده و امکان انتقال مقادیر نسبتاْ زیادی از مواد ژنتیکی را دارد. در مهندسی گیاهی، عناصر همیوغی شبه آگروباکتریوم، مکمل سایر سیستمهای انتقال دهنده استاندارد مانند ویروس موزاییک توتون (tobacco mosaic virus یا به اختصار TMV) میباشد. در حالی که TMV قادر به آلوده کردن بسیاری از خانوادههای گیاهی دولپهای علفی میباشد، همیوغی شبه آگروباکتریوم قادر به آلوده کردن گیاهان دو لپهای و تک لپهای میباشد.
منابع: